




The camera versus the human eye
Superficially, the human eye appears to be similar to a camera. Here looks certainly deceive because, other than a few shared components, the two differ both in operation and in purpose. Consider first the camera. A camera records a scene all at once and in an instant. What is actually recorded is a projection of the scene by a rectilinear or fisheye lens onto a flat sensor. Furthermore, the modern digital camera and the associated post processing computer algorithms are designed to:
Provide maximum resolution with an adjustable depth of field.
Provide enough brightness to reveal some detail in shadows while maintaining detail in bright areas.
Match the color perception of the standard human observer by sampling the image at different wavelengths.
When successful, this setup yields a simulation of standard human vision that stands up to inspection in any part on the image. Each part of the camera’s lens, sensor, and computer is optimized in an effort to match as much as possible the capabilities of the human eye/brain system. The post processing of camera images also attempts to replicate human acutance and color perception. This is, of course, all based on a standard observer.
In contrast to the camera, our eye/brain system scans a scene in many jumps called saccades and processes the incoming stream of information to capture what is important for our well-being. About 2/3’s of the human brain is devoted to processing visual information and its interactions with other senses. Dozens of visual field maps in the visual cortex are involved in the extraction of information concerning motion in various directions, spatial frequencies, recognition of faces, discrimination of light stimuli from random noise of the photo receptors, separation of motion in the surroundings from eye and body motion, and so on.1 The stimuli are processed for immediate action but also to present a coherent picture to the conscious mind. This involves assigning colors to objects, insuring stability of the presented scenes, enabling depth perception, and extrapolating motion so that the time delay in processing will not interfere with conscious actions. All of this amounts to the creation of an amazing simulation of the surrounding world that is based on limited information obtained by an imperfect visual system.
Psychologists have emphasized that we see with our brain using input from the visual cortex, but that our visual images are so processed that they have little direct correlation with images from our eyes. In fact, the brain can produce perfectly realistic scenes even when our eyes are closed. This is, of course, the origin of dreams and hallucinations.
Do our eyes even permit us to perceive the same colors?
The obvious question is “do we all experience the same visual image when confronted with a scene?” If our eye/brain systems are essentially identical, it is still an unanswerable question. When I say that an object is red, how do I know that you are experiencing the same color when you agree that it is red? An easier question to answer is whether you and I can distinguish the same pairs of colors. For example experiments have been designed to determine whether two observers will select the same amounts of red and green light in a mixture to match a given shade of yellow light.
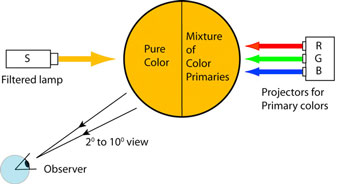
All photographic color matching systems incorporate the idea of a standard human observer. The standard observer is just the average of a number of observers who match colored lights in an experiment. The original standard observer was introduced in the 1931 by the International Commission on Illumination (CIE). A group of about 20 individuals compared a screen illuminated with a pure color, i.e. a spectral color with a known wavelength, with a screen illuminated by the overlapping beams of red, blue, and green light from a selected RGB standard set. The observers adjusted the intensities of the three primaries until a match was achieved with the light from the test lamp. The 1931 experiment used a two degree field of view to just cover the human fovea, while the 1964 CIE supplementary standard expanded the field of view to 10 degrees to cover part of the periphery as well (Fig. 1).

The CIE standard observer is a reasonable choice for designers and photographers, but unfortunately many people are not exactly like the standard observer. Consider the structure of the human eye. All vision depends on our retina, the layer of tissue on the inside back surface of the eye. We have in common a set of about 125 million rod cells in our retina. These photo receptors have maximum sensitivity at about 498 nm, and they are responsible for our night vision. An individual rod can detect a single photon of light, and our visual system is designed so that we can perceive a pulse of light on the eye with as little as seven photons. However, at high light levels, the rod cells are totally saturated and make no contribution to our vision. This brings us to the light receptive cone cells that are much less sensitive and can function in normal daylight. There are six to seven million cones and they are responsible for all human color vision. The cones are concentrated in the foveola, a disk less than 0.3mm in diameter, arranged so that they can also provide high resolution vision as well. It is in the cones that the differences in human color vision are found.
What is color?
Color is not a topic in physics. Pure, monochromatic, light is characterized by its intensity and wavelength. The combination of pure light from two different sources simply gives a light mixture with two different wavelengths. To the standard observer, the combination of the two sources will be perceived as a single color because of the processing of stimuli from cone cells in the retina and the visual cortex. For example a screen illuminated by yellow and blue lights will appear white or near white. Similarly, a red and green combination will be perceived as yellow. Color is assigned to an object by the brain on the basis of the wavelengths of both the object and its environment. But the full set of colors perceived by the standard observer requires that retina contain three types of cones that are sensitive to different ranges of wavelengths. We say that the human eye contains red, green, and blue cones, which is just shorthand for long wavelength (L), medium wavelength (M), and short wavelength (S) receptors, respectively.
The presence of red, green, and blue cones in human eyes and the structures of the cones themselves have now been established by many experiments, but their existence was not at all obvious when Isaac Newton demonstrated the separation of a light beam into its component wavelengths (colors) in 1664. The idea that there are different kinds of photo receptors in the retina of the human eye was presented by the English polymath, Thomas Young in 1801, and he hit the nail on the head by proposing that there are just three types without any experimental evidence to back up his idea. He knew that three colors of light seemed to be sufficient to produce the full array of colors perceived by humans, but he had no way to know that there were not four or more different types of receptors.
In fact, mammals can have from zero to three types of cones.3 Nocturnal animals have up to 100% rods to enhance night (scotopic) vision and thus experience monochromatic vision. Owl monkeys and marine mammals have only one type of cone and are called cone monochromats. New world male monkeys have two types of cones and 60% of females have three types, while old world monkeys and humans share the trichromic visual system. Reptiles and amphibians are usually trichromats as well, but birds and tropical fish have more complex color vision than humans. Apparently, mantis shrimp hold the record with up to 12 types of spectral receptors. Furthermore, in honey- and bumblebees we even find a visual range that extends into the ultraviolet.
This excursion into the animal kingdom was intended to show that the perception of color must vary wildly, and it is just a construct of the eye-brain system. With humans we also find a range of different cone sets that include standard observers as well as a variety of types of color blindness and color anomaly. Anomaly can involve hybrid genes that produce subtle deviations from standard trichromatic vision. In past centuries such deviations would have gone unnoticed, but now with color management methods in photography we can detect subtle differences in color matching abilities.
Now let’s take a brief look at the human system of cones.3 The red, green, and blue visual pigments are all opsin molecules that contain the chromaphore 11-cis-retinal but differ slightly in their amino acid sequences. Amino acid substitutions in the opsin chains tune the aborption spectra of the chromaphores and account for the wavelength shifts. The genes that express the red and green pigments are located on the X chromosome while the gene for the blue pigment is on chromosome 7. Figure 2 shows the absorption spectra for the red (L), green (M), and blue (S) pigments as well as for the rods (dotted line). The pigments that are responsible for red and green absorptions have quite similar amino acid sequences and are said to be highly homologous. This accounts for the similar spectra and the small wavelength shift between them. Furthermore the genes for red and green are located side by side on the DNA of the X-chromosome with red coming first. The arrangement LM is normal but usually there are two or more copies of the M gene, and this causes no problems.

Genetics of color vision anomaly
The most common kinds of color vision defects occur when a red (L) or green (M) gene is completely missing.5 A protanope is a person who is missing the L gene and a deuteranope is missing the M gene. In either case the individual cannot distinguish greens and reds. A simulation of these forms of dichromatic vision is presented in Fig. 3. Trianopes, who are quite rare, are missing the S gene.

The loss of an L or M gene usually occurs by means of gene recombination that can occur during mitosis. For example, two DNA chains with the LM combination interact to produce one chain with only L and one with LMM as illustrated in Fig. 4. A male receiving an X-chromosome with only an L gene is sure to be a deuteranope. Females, of course have two X-chromosomes which are randomly expressed so that a missing gene on one X-chromosome can be made up for by the expression of the other X-chromosome.

This is just the beginning of the possible types of red green anomalous vision. The most common variations in trichromats result from the presence of hybrid genes. The hybrids result from gene recombinations that produce red genes with some green component (L/M), deuteranomaly, or green genes with some red component (M/R), protanomaly, as shown in Fig. 5. All of the red/green vision anomalies amount to 8% to 10% of human X-chromosomes and hence the same percentages men with anomalous vision. Women, whose risks are cut by having two X-chromosomes, have only about 0.5% chance of anomalous color vision.

As a final example of visual diversity I would like to consider a difference between “normal” trichromats that is revealed in color matching experiments. Two types of L genes that differ in a single nucleotide are found on X-chromosomes. This results in L photopigments that have either serine (ser) or alanine (ala) at position 180. The absorption spectrum of pigments with ser are red shifted by 4 nm to 5 nm. Males, having only one X-chromosome, have either the ser or the ala version of the L gene while women can have either or both in the case of heterozygotes. In color matching experiments to determine the amounts of red and green light require to match a given standard yellow light, some males require more red than others. This results in a bimodal distribution for males while females exhibit either the matching selections shown by males or something in between for heterozygotes. Therefore, some women have four cones and exhibit tetrachromacy. Humantetrachromats potentially can distinguish colors better than trichromats.
Dealing with differences in color perception
This discussion has shown that 8-10% of human X-chromosomes encode color vision anomalies, and that there are even differences in color perception of “normal” trichromats. I find it strange that these differences are seldom mentioned in discussions of color management and color correction for photographs. Of course, photographers want to have their photographs critiqued and may want to sell them; so the opinions of “standard observers” are very important. Presumably, protanopes and deuteranopes are aware of their condition and have been dealing with it. But those with other more subtle anomalies may be unaware how much they deviate from “normal” trichromates in color perception. This is made more complicated by the metameric nature of color matching, that is to say the same perceived color can be matched by combinations of different sets of primary colors. Even in the group of “normal” trichromats there are differences in color matching abilities that may result from different pigment densities in the retina or from age related changes in eyes, e.g. cataract related lens color.
Most differences in the ability to discriminate colors are permanent, and we should take this into account when we produce images that will be viewed by others. This applies both to those with deviations in color perception and those who create photographs and illustrations for a wide audience including those with a wide range of color responses. Consider the following situations:
Obvious red/green color blindness: This condition is found in up to about 5% of the population, and these dichromats must adapt to color standards set by standard observers who are trichromats. In the post processing of images, this does not have to be a major problem. Small enhancements of saturation, clarity, and vibrance can be made without reference to red/green differences, and of course photographs can be critiqued by friends.
Subtle differences in the group of trichromats: Groups of individuals who behave differently in color matching experiments are said to be metameric observers. More differences may be found when light sources, the color primaries, are changed; and this brings us to the topic of modern display technologies.6 Some individuals may be happy with color presentation on CRT displays but not with high gamut LCD screens. In Figure 6 the spectral distribution for a CRT type display is shown. The blue and green bands are very wide and the red bands are scattered over a wide range. These distributions can mask differences shifts in the absorption maxima. In contract to this, the RGB band widths may be quite small in LCD displays as shown in Fig. 7. Accordingly, LED monitors may accentuate distinguishing shifts in the absorption spectra of anomalous photo pigments.


Photographers, designers, and illustrators: This group needs to be sensitive to the ways their audiences will see the work they create. There are various aids to help avoid confusion and even danger for color blind individuals. For example the EIZO ColorEdge® monitor simulates the way color schemes for web sites, signs, maps, etc., will be viewed by those with red-green (protanopia and deuteranopia) and blue-yellow (tritanopia) color vision deficiencies.
Summary
Our eye/brain system assigns color to objects in the images it creates, and our perception of color is unique. Our ability to distinguish colors depends on the set of cones we inherit.
Mammalian eyes contain rods and up to three types of cones that enable color vision. Standard human observers are trichromats with three types of cones.
Up to 10% of males and about 0.5% of females have anomalous vision. Red/green vision problems resulting from deficiencies in an X-chromosome are the most common. Females have two X-chromosomes and this provided backup.
Normal human long wavelength (L or red) cones are divided into two groups because of a single amino acid substitution in the associated photo pigment. This results in a small red shift in the absorption maximum of the L cone for half of the male population. Because females have two X-chromosomes they may inherit either form of the gene or both. Therefore, many females are tetrachromats!
Those having anomalous vision must adapt to color standards set by standard observers who are trichromats. However, all designers and photographers should take into account the way their creations will be viewed by those with various color anomalies. Non-standard color vision can be simulated in photo processing software such as Photoshop® and in some color monitors.
The narrow band primary colors used in LCD displays may have the effect of amplifying small color shifts in photo pigments, and some people may detect differences in color matches with CRTs relative to LCDs.
References and Links
Dale Purves and R. Beau Lotto (2011), Why We See What We Do Redux, A Wholly Empirical Theory of Vision, Sunderland, MA: Sinauer Assoc., Inc. p. 214.
Charles S. Johnson, Jr. (2010), Science for the Curious Photographer, New York, NY: CRC Press.
Strongly recommended neuroscience lectures by Jeremy Nathans: http://www.ibiology.org/ibioseminars/neuroscience/jeremy-nathans-part-1a.html
Bowmaker J.K. and Dartnall H.J.A., J. Physiol. 298: pp501-511 (1980), Wikipedia Commons
Samir S Deeb, PhD and Arno G Motulsky, MD., “Red-Green Color Vision Defects, Red-Green Color Blindness,” http://www.ncbi.nlm.nih.gov/books/NBK1301/
Abhijit Sarkar, Laurent Blondé, Patrick Le Callet, Florent Autrusseau, Patrick Morvan, Jürgen Stauder, “Toward Reducing Observer Metamerism in Industrial Applications: Colorimetric Observer Categories and Observer Classification,” 18th Color Imaging Conference Final Program and Proceedings, Society for Imaging Science and Technology, 2010.
About Charles Johnson
I was trained in Physical Chemistry at MIT, and I am now Professor Emeritus of Chemistry at the University of North Carolina in Chapel Hill. I am also an avid nature photographer, and I blog about photography at www.photophys.com. My scientific background is very helpful to me as I research and write about the physical and psychological bases of photography. My book, Science for the Curious Photographer, was published by A K Peters/CRC in 2010.
© Charles S. Johnson, Jr., 2014
Published December 12, 2014





Elevate Your Vision



Read this story and all the best stories on The Luminous Landscape
The author has made this story available to Luminous Landscape members only. Upgrade to get instant access to this story and other benefits available only to members.
Why choose us?
Luminous-Landscape is a membership site. Our website contains over 5300 articles on almost every topic, camera, lens and printer you can imagine. Our membership model is simple, just $2 a month ($24.00 USD a year). This $24 gains you access to a wealth of information including all our past and future video tutorials on such topics as Lightroom, Capture One, Printing, file management and dozens of interviews and travel videos.
- New Articles every few days
- All original content found nowhere else on the web
- No Pop Up Google Sense ads – Our advertisers are photo related
- Download/stream video to any device
- NEW videos monthly
- Top well-known photographer contributors
- Posts from industry leaders
- Speciality Photography Workshops
- Mobile device scalable
- Exclusive video interviews
- Special vendor offers for members
- Hands On Product reviews
- FREE – User Forum. One of the most read user forums on the internet
- Access to our community Buy and Sell pages; for members only.
You may also like



